Rats can't vomit. They can't burp either, and they don't experience heartburn. Rats can't vomit for several related reasons: (1) Rats have a powerful barrier between the stomach and the esophagus. They don't have the esophageal muscle strength to overcome and open this barrier by force, which is necessary for vomiting. (2) Vomiting requires that the two muscles of the diaphragm contract independently, but rats give no evidence of being able to dissociate the activity of these two muscles. (3) Rats don't have the complex neural connections within the brain stem and between brain stem and viscera that coordinate the many muscles involved in vomiting.
One of the main functions of vomiting is to purge the body of toxic substances. Rats can't vomit, but they do have other strategies to defend themselves against toxins. One strategy is super-sensitive food-avoidance learning. When rats discover a new food, they taste a little of it, and if it makes them sick they scrupulously avoid that food in the future, using their acute senses of smell and taste. Another strategy is pica, the consumption of non-food materials (particularly clay), in response to nausea. Clay binds some toxins in the stomach, which helps dilute the toxin's effect on the rat's body.
Vomiting, or emesis, is the reflexive act of ejecting stomach contents forcefully through the mouth by coordinated muscular contractions.
Why do humans vomit?
One of the main functions of vomiting is to rid the body of toxic substances. The body has several hierarchical lines of defense against toxins (Davis et al. 1986):
- First line of defense: Avoidance of certain foods due to smell or taste cues
- Second line of defense: Detection of toxins in the gut followed by nausea (prevents further consumption) and vomiting (purges the body of already ingested toxin)
- Third line of defense: Detection of toxins in the circulation by a sensor in the central nervous system, also followed by vomiting.
Mechanisms of human vomiting
Vomiting is a complex suite of coordinated muscular actions, controlled by a group of nuclei in the brainstem. In essence, great pressure is put on the stomach by surrounding muscles and the esophagus is opened. The result is that the stomach's contents are expelled forcefully from the mouth (Fig 1).
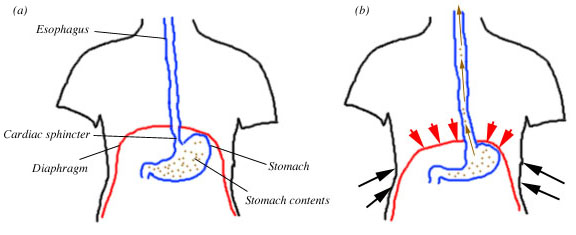
Figure 1. The emetic reflex in humans. (a) digestive system at rest, and (b) digestive system during the emetic reflex. The diaphragm puts pressure on the stomach, the esophagus opens, and the stomach contents are expelled forcefully into the esophagus and out of the mouth.
More specifically, during vomiting the muscles of the abdomen and chest contract and the diaphragm spasms downward and inward, which all put pressure on the stomach. In the next phase, the part of the diaphragm that surrounds the esophagus relaxes, thus helping to open the esophagus. The longitudinal muscle of the esophagus contracts, further opening the junction between the stomach and esophagus. The pressure forces the contents of the stomach up into the esophagus and out of the mouth (for reviews with much more detail, see Brizzee 1990, Lang and Sarna 1989, Miller 1999).
Rats and vomiting
Rats are considered a non-vomiting species (also called nonemetic) (Hatcher 1924). Rats do not vomit in response to cues that cause vomiting in other animals, like emetic drugs, poison, motion-sickness, and radiation (e.g. Takeda et al. 1993). Rats also don't belch and experience hardly any reflux (heartburn).
Regurgitation vs. vomiting
Rats cannot vomit, but they do regurgitate occasionally. Regurgitation is different from vomiting. Vomiting is the forceful expulsion of stomach contents from the mouth. Vomiting is an active process: it is a complex, powerful reflex requiring the coordination of many muscles. In contrast, regurgitation is the passive, effortless flow of undigested stomach contents back into the esophagus. Regurgitation happens without any forceful abdominal contractions.
There is at least one report of rats choking on regurgitated stomach contents (Will et al. 1979). Upon necropsy, the regurgitated stomach contents (regurgitant) were found to be thick and pasty. They were packed into the rats' pharynx, larynx and esophagus. The action of the tongue had packed the regurgitant into a plug, causing choking. The rats' tongues were also lacerated or bruised from attempts to remove the material by chewing or clawing. Regurgitation was more common in rats fed bulky diets than those fed on standard diets, and more common in females than in males.
Other actions that may resemble vomiting, but are not
Difficulty swallowing, choking: Rats may have trouble swallowing a food item. A rat who has trouble swallowing a food item may strain intently, pull his chin down toward his throat and flatten his ears. He may drool saliva, paw at his mouth, and rub his mouth on nearby surfaces. Most rats are still able to breathe through this (true choking is rare in rats), and work the food out themselves in time, but serious cases may require veterinary asssitance.
Difficulty swallowing may superficially resemble vomiting because partly processed food may come back out of the mouth, but it is not vomiting, which is the forceful, rapid, coordinated, reflexive explusion of stomach contents.
Respiratory distress: rats may be found choking, gagging on, or struggling to breath through a cream or tan colored foamy substance. This foam is not made of stomach contents, but of mucus brought up from the lungs that has been whipped up into a froth. This foam is a symptom of a respiratory problem, not regurgitation or vomiting (pers comm B. Mell D.V.M., 2004).
Rat stomach anatomy
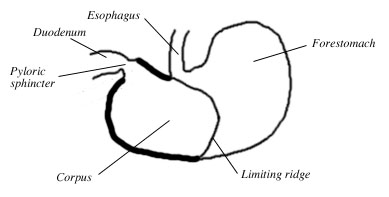
Figure 2. Diagram of the rat's stomach. Adapted from Moore 2000.
The rat's stomach has two parts (Robert 1971):- Forestomach: thin-walled, non-glandular section that receives the esophagus and serves as a holding chamber for food. Its walls are similar to those of the esophagus.
- Corpus: thick-walled, glandular section. Its walls have secretory glands that produce digestive enzymes
and mucus. Digestion begins in the corpus. The pyloric sphincter controls the movement of food from the corpus to
the intestines (specifically, the duodenum).
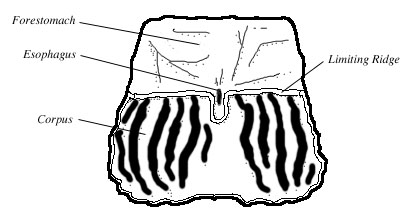
Figure 3. Diagram of a rat stomach opened along the greater curvature of the stomach. Adapted from Robert 1971.
The forestomach and the corpus are separated by a low fold of tissue called the limiting ridge (margo plicatus). The limiting ridge extends circumferentially from the large curvature of the stomach to the small curvature, just below the esophagus. At the esophagus, the course of the limiting ridge bends into a U-shape and almost surrounds the esophageal opening (Luciano and Reale 1991, Robert 1971, Botha 1958) (Fig 3).
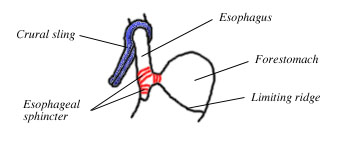
Figure 4. Diagram of the crural sling and the muscle bundles of the esophageal sphincter, which make up the gastroesophageal barrier and are responsible for closing the esophagus. Adapted from Montedonico et al. 1999a.
The rat's esophagus has two layers of striated muscle (outer longitudinal and inner circular), which become smooth near the attachment point with the stomach. The esophagus is closed off from the stomach by the gastroesophageal barrier, which consists of the crural sling, the lower esophageal sphincter, and the several centimeters of intraabdominal esophagus that lie between them (Soto et al. 1997; Fig 4). Humans also have a crural sling and an esophageal sphincter, but ours are placed right on top of one another (Mittal 1993). In rats, they are separated by several centimeters of intraabdominal esophagus (Soto et al. 1997.)
The crural sling is part of the diaphragm (its outer contour is continuous with the diaphragm). It is a U-shaped bundle of fibers that wraps around the esophagus and attaches to the vertebrae. When the crural sling contracts it pinches the esophagus closed.
The esophageal sphincter is a circular muscle that surrounds the base of the esophagus. At its lower edge, it has muscle fibers that insert into the limiting ridge (Fig 4). So when the sphincter contracts, it not only constricts the walls of the esophagus, it also pulls the sides of the limiting ridge's "U" together, thus hiding and tightly closing the esophageal opening (Montedonico et al. 1999b, McKirdy and Marshall 2001, Botha 1958) (Fig 5).
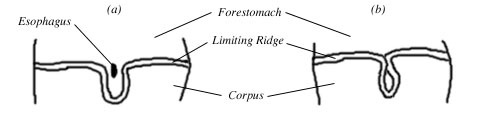
Figure 5. Diagram of the limiting ridge and the esophageal opening in the rat's stomach when the esophageal spincter is (a) open and (b) closed. Adapted from Montedonico et al. (1999b).
How rats can't vomit
Anatomical textbooks on rats usually mention in passing that rats can't vomit. They tend to implicate the limiting ridge or the lack of striated muscle in the rat's esophagus, and sometimes both (Fox et al. 2002, Haschek and Rosseaux 1998, Moore 2000, Rice and Fish 1994, Turton et al. 1998), but these textbooks do not go into more detail about exactly how these features of a rat's anatomy prevent a rat from vomiting, or if there are any other features involved.
Looking deeper into the scientific literature, I found a complex story about why a rat is unable to vomit:
Rats have a powerful and effective gastroesophageal barrier, consisting of the crural sling, the esophageal sphincter, and the centimeters of intraabdominal esophagus (see above). The pressure at the two ends of this barrier is much higher than the pressure found in the thorax or abdomen during any phase of the the breathing cycle (Montedonico et al. 1999b). The strength and pressure of this barrier make reflux in rats nearly impossible under normal conditions (Montedonico et al. 1999a), though Will et al. (1979) records low rates of regurgitation.
In order to vomit, the rat would have to overcome this powerful barrier. Evidence suggests that rats cannot do this, because (1) they can't open the crural sling at the right time, and (2) they can't wrench open the esophageal sphincter. In addition, (3) rats lack the necessary neural connections to coordinate the muscles involved in vomiting.
(1) Rats can't relax the crural sling while contracting the rest of the diaphragm. The diaphragm is has two muscles: the crural (muscle fibers attached to the vertebrae, called the crural sling) and the costal (muscle fibers attached to the rib cage). The esophagus passes through the crural sling, so when the crural diaphragm contracts the esophagus is pinched closed.
During the expulsive phase of vomiting in humans, the activity of these two diaphragm muscles diverges. The costal section contracts, putting pressure on the stomach, while the crural section relaxes, allowing stomach contents to pass through the esophagus (reviewed in Pickering and Jones 2002). Rats, however, do not dissociate the activity of these two parts of their diaphragm: they do not relax the crural section while contracting the costal section. Instead, both muscles contract or relax together (Pollard et al. 1985). The rat's inability to separately and selectively control its two diaphragmatic muscles therefore plays an important role in its inability to vomit: the rat can't put the necessary pressure on the stomach and open the crural sling to allow the contents to escape at the same time.
(2) Rats can't wrench open their esophageal sphincter. In humans, the esophageal sphincter is opened during vomiting with the help of the longitudinal muscle of the esophagus (Lang and Sarna 1989). This allows the expulsion of stomach contents during vomiting. Rats, however, have only a thin, weak longitudinal muscle which is unstriated where it joins the stomach. It is too weak to wrench open the sphincter and permit the evacuation of stomach contents (Steinnon 1997).
(3) Rats lack the necessary neural connections within the brain and between brain and viscera. Animal species that vomit have a "vomiting center" in the brainstem, consisting of several interconnected nuclei that coordinate all the many muscles involved in vomiting (see Borison and Wang 1953). Animals that don't vomit, like rats and rabbits, have the brainstem nuclei and the muscle systems used in vomiting, but they don't have the complex connections between the nuclei or between the brainstem and the viscera that are required for such a coordinated behavior (King 1990).
Why rats can't vomit, and what they do instead
Is it beneficial for rats to be unable to vomit?
As of yet, no empirical research has been done on whether the inability to vomit benefits the rat in some way. Davis et al. (1986) provides some interesting speculation on this topic, however. Remember that Davis et al. suggested that there are hierarchical lines of defense against toxins (first food avoidance, then detection of toxins in gut, and lastly detection of toxins in circulation, followed by vomiting). Davis et al. notes that rats have extremely sensitive senses of smell and taste (Roper 1984). The rat uses its senses of smell and taste to avoid foods that made it feel ill in the past (Garcia et al. 1966, Rozin and Kalat 1971). In fact, rats avoid foods in response to cues that cause vomiting in other species (Coil and Norgren 1981). So the rat who avoids foods that made it feel ill should not ingest lethal amounts of that food in the future.
Davis et al. speculates that because rats have such an extraordinarily well-developed first line of defense against toxins (conditioned food avoidance), the rats' later lines of defense (vomiting in response to gastric or circulatory cues) have become redundant and were therefore lost over time. Rats can, in fact, detect toxins in the stomach (Clarke and Davison 1978), and in the circulation (Coil and Norgren 1981) but they don't respond by vomiting, instead they avoid that food in the future. So, the theory goes, rats have lost the ability to vomit because they no longer need it: rats never eat lethal amounts of toxic foods in the first place.
However, an alternative theory is that rats developed their hyper-sensitive food avoidance to compensate for the inability to vomit. It makes sense for a rat to scrupulously avoid ingesting toxic food if it can't get rid of it later. So, it might indeed benefit the rat to be able to vomit, but as vomiting isn't an anatomical option, the rat has developed other methods of protecting itself, including food avoidance.
Also, rats do still need a strategy to cope with ingested toxins. Rat food avoidance isn't foolproof. Rats do experience nausea and have evolved an alternative to vomiting: pica, the consumption of non-nutritive substances. When rats feel nauseous they eat things like clay, kaolin (a type of clay), dirt and even hardwood bedding (eating clay and dirt is a type of pica called geophagia). Their consumption isn't random, though: rats offered a mixture of pebbles, soil and clay after being given poison prefer to eat the clay (Mitchell 1976).
Rats engage in pica in response to motion-sickness (Mitchell et al. 1977a, b, Morita et al. 1988b), nausea-inducing drugs (Mitchell et al. 1977c, Clark et al. 1997), radiation (Yamamoto et al. 2002b), and after consuming poisons (Mitchell 1976), or emetic drugs (Takeda et al. 1993). The incidence of pica decreases in response to anti-emetics (Takeda et al. 1993) and anti-motion sickness drugs (Morita et al. 1988a). Pica in rats is therefore analogous to vomiting in other species.
The consumption of non-nutritive substances may be an adaptive response to nausea. Nausea is frequently caused by a toxin, and non-nutritive substances may help dilute the toxin's effect on the body. Clay in particular binds and inactivates many types of chemicals and is therefore good at deactivating toxins (e.g. Philips et al. 1995, Philips 1999, Sarr et al. 1995). Pica may therefore be part of the rat's second line of defense against toxins.
Evolution of vomiting
The evolution of behavior is studied by examining and comparing the behavior of living species (see Martins 1996 for more). The behavior is then mapped onto the phylogeny, or evolutionary "family tree" of those species, and deductions can then be made about when a particular behavioral trait appeared in the past. For example, if a group of related species exhibit the same behavior, then their common ancestor probably did, too. If just one species in a group has a particular behavior, then that behavior probably wasn't present in the common ancestor, but evolved later just in that species.
Such broad comparative studies involving dozens of species have not yet been done for vomiting. In fact, it is a bit difficult to determine how common vomiting is in the animal kingdom: Hatcher (1924) says that the ability to vomit is a primitive, common trait and many species do it. Harding (1990), however, states that very few species are capable of vomiting. Until a survey of many different species is done, we won't know the answer for sure.
A survey of the literature shows that information on vomiting does exist for a few species (Table 1).
|
Species |
Vomits? |
Species differences |
Reference |
|
Mammals |
|||
|
Primates |
|||
| Human |
Yes |
responds to all known emetics |
Borison 1981 |
| Rhesus monkey (Macaca mulatta) |
Yes |
- |
Borison and Wang 1953 |
| Crab-eating macaque (M. fascicularis or cynomologus) |
Yes |
- |
Borison and Wang 1953 |
| Squirrel monkey |
Yes |
- |
Brizzee et al. 1980, Ordy and Brizzee 1980 |
|
Carnivores |
|||
| Dog |
Yes |
responds to all known emetics |
Borison 1981, Noble 1945 |
| Cat |
Yes |
responds to most emetics |
Borison 1981, Noble 1945 |
| Ferret |
Yes |
responds to most emetics |
Knox et al. 1993, Florczyk et al. 1982 |
|
Even-toed ungulates |
|||
| Pig |
Yes |
responds to selected emetics |
Borison 1981 |
| Sheep |
Yes |
vomits rarely |
Borison 1981 |
|
Odd-toed ungulates |
|||
| Horse |
Yes |
vomits rarely, if ever |
Borison 1981 |
|
Cetaceans |
|||
| Sperm whale |
Yes |
yes |
Clark et al. 1988 |
|
Rodents |
|||
| Norway rat |
No |
never vomits |
Borison 1981 |
| House mouse |
No |
never vomits |
Yamamoto et al. 2002 |
| Woodchuck |
Yes |
may vomit from Red Squill poison |
Borison 1981 |
| Guinea pig |
No |
never vomits |
Ossenkopp and Ossenkopp 1985, 1990 |
|
Lagomorphs |
|||
| Rabbit |
No |
never vomits |
King 1990 |
|
Insectivores |
|||
| Least shrew (Cryptotis parva) |
Yes |
- |
Darmani 1998 |
| Asian musk shrew (Suncus murinus) |
Yes |
responds to selected emetics, doesn't have CTZ (brain area involved in vomiting) |
Ueno et al. 1987 |
|
Birds |
|||
|
Galliformes |
|||
| Chicken |
Yes |
- |
Yang et al. 1999 |
| Japanese quail |
No |
- |
Ossenkopp & Tu 1984 |
|
Collumbiformes |
|||
| Pigeon |
Yes |
- |
King 1990, Saxena et al. 1977 |
|
Procellariiformes |
|||
| European storm petrel (Hydrobates pelagicus) |
Yes |
- |
Matthews 1949 |
|
Reptiles |
|||
|
Crocodiles |
|||
| Estuarine crocodile (Crocodylus porosus) |
Yes |
- |
Andrews et al. 2000 |
|
Amphibians |
|||
|
Anurans (frogs) |
|||
| African clawed frog (Xenopus laevis) |
Yes |
- |
Naitoh et al. 2000 |
| Schlegel's green tree frog (Rhacophorus schlegelii) |
Yes |
- |
Naitoh et al. 2000 |
| Wrinkled frog (Rana rugosa) |
Yes |
vomiting response varies seasonally |
Naitoh et al. 2000 |
| Black-spotted pond frog (Rana nigromaculata) |
Yes |
- |
Wassersug et al. 1993 |
| Japanese frog (Rana porosa porosa) |
Yes |
- |
Miyachi et al. 2002 |
| Japanese tree frog (Hyla japonica) |
Yes |
- |
Wassersug et al. 1993 |
|
Urodeles (salamanders) |
|||
| Clouded salamander (Hynobius nebulosus) |
Yes |
- |
Wassersug et al. 1993 |
|
Fish (Osteichthyes: bony fish) |
|||
|
Salmoniformes |
|||
| rainbow trout (Oncorhynchus mykiss) |
Yes |
- |
Tiersch and Griffith 1988 |
|
Perciformes (perch-like fish) |
|||
| bluefin tuna (Thunnus thynnus) |
Yes |
- |
Carey et al. 1984 |
|
Fish (Actinopterygii: ray-finned fishes) |
|||
|
Siluriformes (catfishes) |
|||
| African walking catfish (Clarias gariepinus) |
Yes |
emetic reflex develops with age |
Sims and Andrews 1996 |
|
Chondrichthyes: rays, sharks, and relatives |
|||
|
Carcharhiniformes (ground sharks) |
|||
| lesser spotted dogfish (Sciliorhinus canicula) |
Yes |
- |
Andrews et al. 1998 |
Tentatively, from this table, it looks like the ability to vomit is widespread among vertebrates, and hence is an evolutionarily old trait that appeared in a distant common vertebrate ancestor and was passed down to its many descendants. The ability to vomit may then have been lost in the common ancestor of rodents and rabbits (and later regained in the woodchuck). Further inquiry into the exact mechanisms of vomiting in different species would cast more light on whether vomiting has evolved multiple times.
Note that these deductions are purely speculative and casual at this point, as they are based on just a few scattered species. A broad, comprehensive survey of many different species is needed before the evolution of vomiting is fully understood.